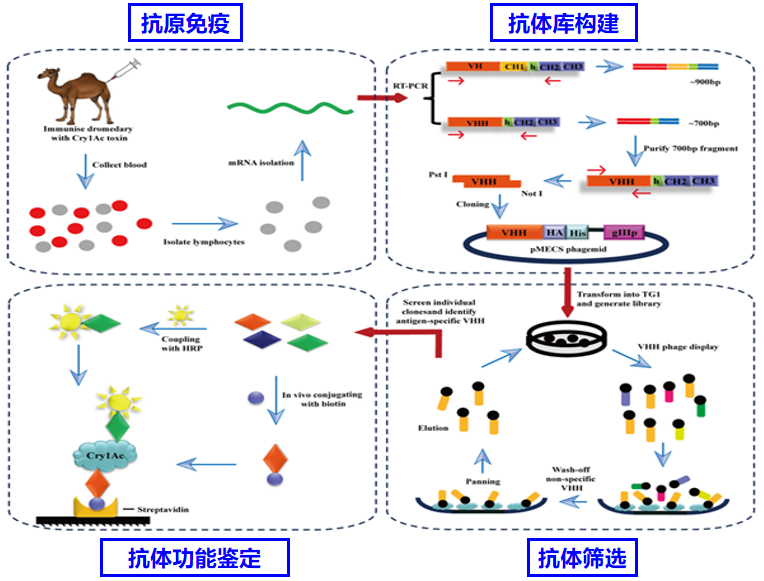
1993年,比利时布鲁塞尔自由大学免疫学家Hamers-Casterman教授以及他的同事们在骆驼(骆驼科,后来研究证实也包括 单峰骆驼和羊驼)的血清中发现了一种与传统抗体结构不同的新型抗体,这种抗体仅仅由两条重链构成,被称为重链抗体(heavy-chain antibody, HCAb)。重链抗体的重链的可变区称为VHH(variable domain of heavy chain of heavy-chain antibody),通过体外重组表达制备的VHH 分子质量仅仅为15kDa, 是传统抗体的十分之一左右,是抗原结合片段(scFV,VH-VL)的二分之一左右,因此被称为纳米抗体(nanobody, Nb)。
由于没有轻链,纳米抗体仅有3个属于重链的抗原识别区域(CDRs 区)。为了弥补缺少的轻链CDRs的生物学活性,纳米抗体在 CDR3部分增加了氨基酸长度(16~18个氨基酸残基,对应的传统抗体的CDR3有8-15个氨基酸),以增加CDR的多样性和特异性。另外,纳米抗体的 CDR3 区域可形成一个大的暴露的凸环,像“手指”一样延伸到抗原的缝隙或者裂口中,可以接触到传统抗体不能接触的抗原表位。CDR3凸环中的一个半胱氨酸还可以与CDR1 或FR2 的 45 位点的半胱氨酸形成二硫键,可使纳米抗体的生物结构相对稳定,大大降低纳米抗体与抗原结合所需能量,使得高亲和力的纳米抗体的获得较容易。